7月 28日,《Nature》新闻特写上刚好发布了一篇关于Delta变异株的长文《How the coronavirus infects cells — and why Delta is so dangerous》
[1]
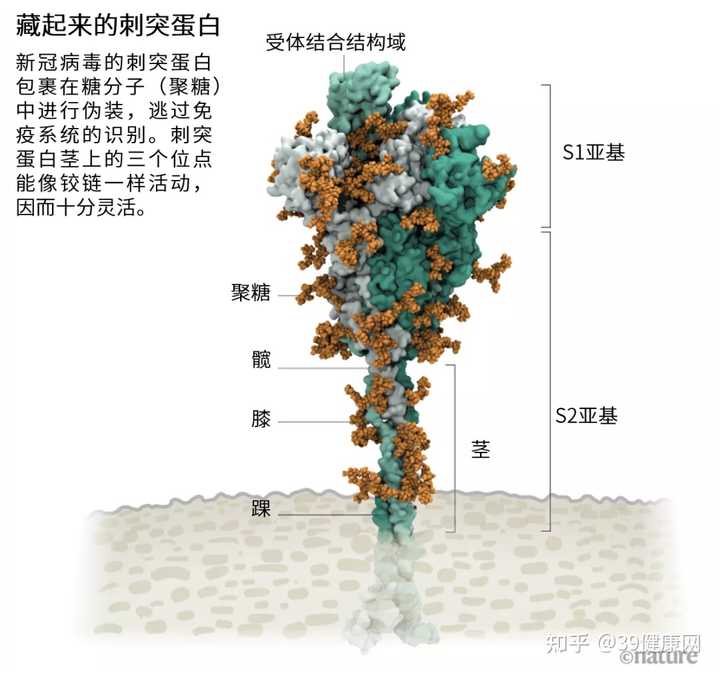
我们都知道,新冠病毒披上了一层奢华的
糖衣——刺突蛋白 。这些刺突蛋白从新冠病毒的表面突起,让病毒包裹在聚糖(glycan)这种糖分子里。
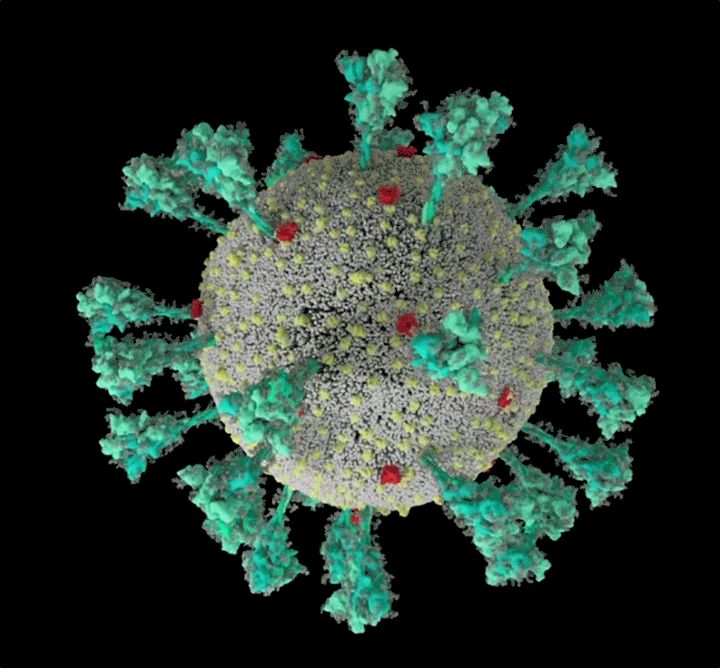
新冠病毒结构的计算机模拟。图源 @Janet Iwasa, University of Utah
许多病毒的外层蛋白质都有聚糖,
就像披着羊皮的狼,逃过人体免疫系统的监视。
而从刺突蛋白顶部突出的
无包裹裸露环状结构是一个受体结合结构域(RBD),是刺突蛋白与人细胞表面受体结合的三个区域之一 (见下图)。
图源 @Structural image from Lorenzo Casalino, Univ. California, San Diego (Ref. 1); Graphic: Nik Spencer/Nature
模型中,
当 RBD上升到这团聚糖的顶端时,两个聚糖会突然将它锁住 ,就像自行车的撑脚架一样。如果改变聚糖,RBD就会倒塌。
研究也证明,改变这两个聚糖会降低刺突蛋白与人细胞受体的结合力,即
剪掉这两个聚糖或能降低新冠病毒的感染性 ,可惜目前还没找到方法。
自新冠大流行以来,科学家们一直在研究了解
新冠病毒是如何感染人体细胞的 。通过分离感染过程,他们希望找到更好的方法,比如通过改进治疗方法和疫苗来中断它,并了解为什么最新的毒株,如 Delta变种会变得更容易传播。
通过 19个月的不懈努力,加之数十年的冠状病毒研究,科学家们正在逐渐解开
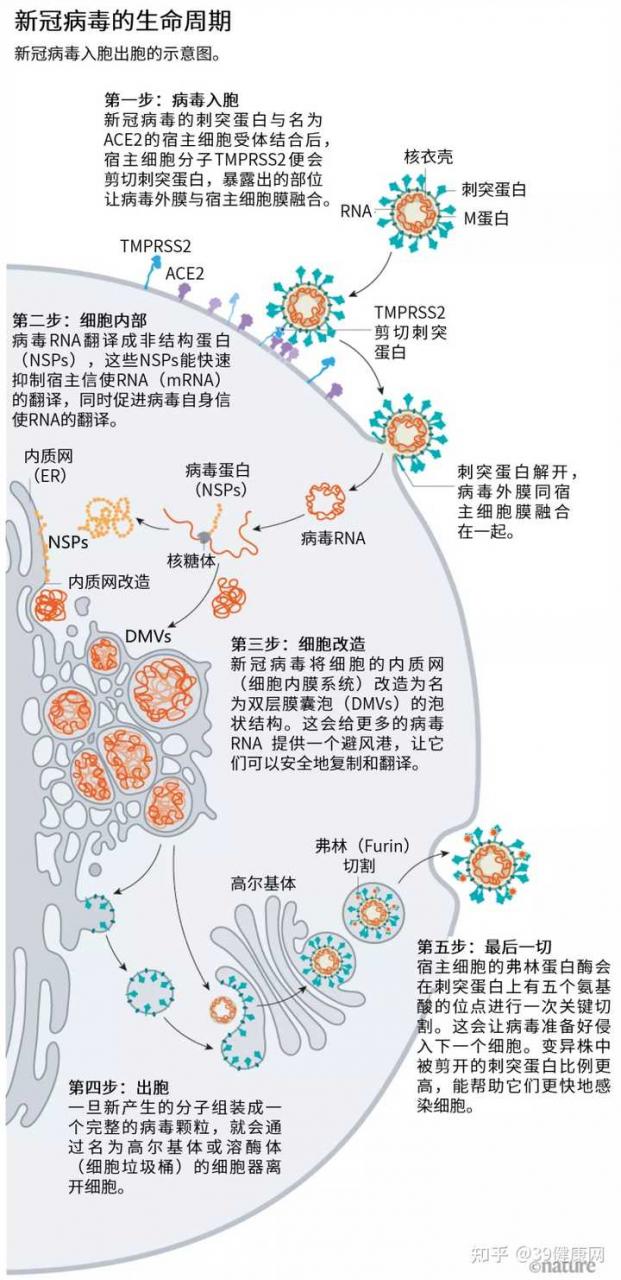
新冠病毒是如何一步步侵入人体细胞的 (见下图)。
来源 @Hui (Ann) Liu, Univ. Utah; 绘图 @Nik Spencer/Nature
研究人员确定了
一种关键适应机制 ,可以帮助新冠病毒以惊人的强度抓住人体细胞,然后在进入人体细胞后将自己隐藏起来。随后,新冠病毒离开细胞时又会使出关键一招,
令其病毒颗粒能继续感染更多人体细胞。
这就是使新冠病毒能够如此
迅速传播 并
夺人性命 的关键,也是新冠病毒
难以遏制 的原因。
步骤① —— 蓄刺待发
一切从刺突蛋白开始。每个新冠病毒颗粒(virion)的表面有
24-40个任意排列的刺突蛋白 ,这是病毒与人类细胞融合的关键。
对于其他类型的病毒,例如流感,外部融合蛋白相对刚性。而新冠病毒的刺突蛋白
非常灵活 ,能在三个点像铰链一样活动。
这使得
刺突蛋白能任意掉头、摇摆、旋转 ,使它们更容易扫描细胞表面,并使多个刺突蛋白与人体细胞相结合。目前其他冠状病毒尚无类似实验数据,但由于刺突蛋白序列在进化上
高度保守 ,可以推测所有冠状病毒都具有这一特点。
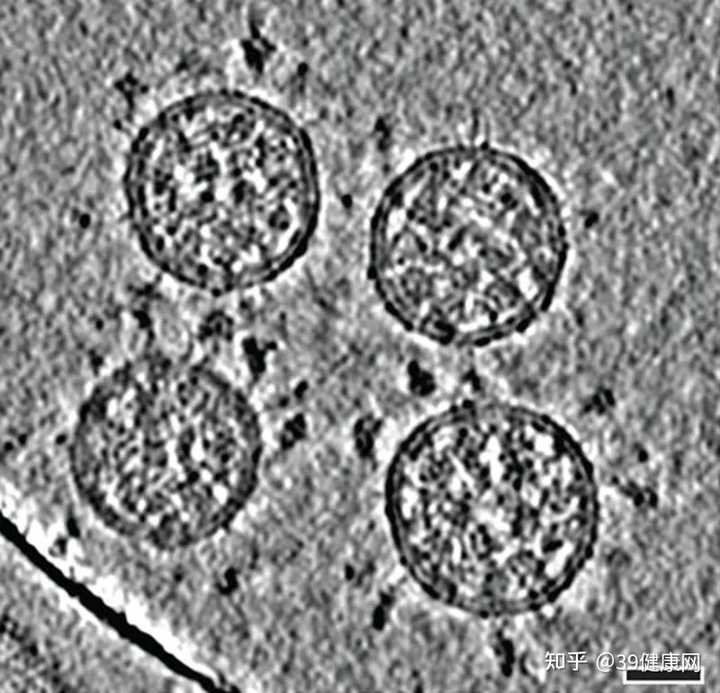
新冠病毒颗粒的冷冻电镜断层扫描成像(比例尺30纳米)图源 @B. Turoňová et al./Science
在疫情早期,研究人员就证实了
新冠病毒刺突蛋白的 RBD能与 ACE2受体结合。
ACE2受体是一种常见的蛋白,广泛分布于人类喉部和肺部的大部分细胞表面。这个受体还是
导致严重急性呼吸系统综合征的 SARS病毒(SARS-CoV)的入胞点。
但是,
新冠病毒与 ACE2的亲和力更好,估计是 SARS病毒的2-4倍 ,这是因为RBD的多种变化能稳定其与病毒结合的热点位置。
而令人担忧的是,新冠病毒变异株往往在刺突蛋白的 S1亚基上发生突变,S1亚基包含 RBD并负责病毒与 ACE2受体的结合。
注:刺突蛋白的 S2亚基则能促进病毒外膜与宿主细胞膜的融合 。
以 Alpha变异株为例,它在刺突蛋白序列上有 10个突变,
导致 RBD更有可能保持【向上 】
的状态,这能让病毒更容易进入细胞。
而 Delta变异株在 S1亚基上有多个突变,包括 RBD上的3个突变。
这 3个突变似乎不仅能提高 RBD与 ACE2的亲和力,还能提高其逃逸免疫系统的能力。 步骤② —— 特定通道
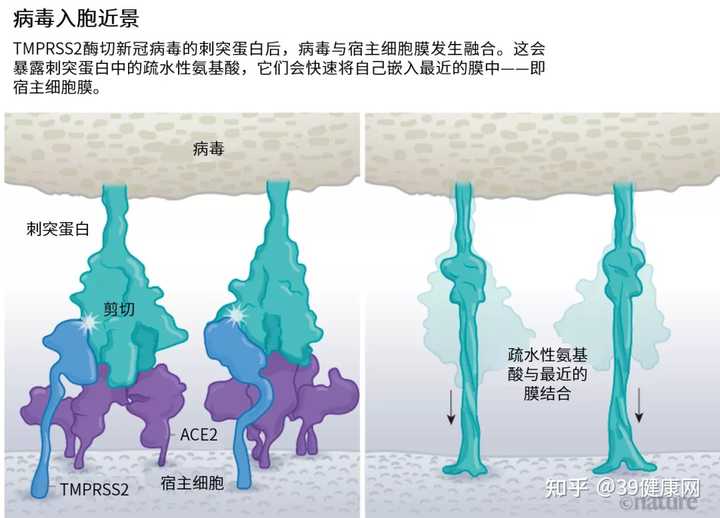
一旦病毒刺突蛋白与 ACE2结合,宿主细胞表面的其他蛋白就会启动一个过程,导致病毒外膜与细胞膜融合 (见下图)。
来源 @Janet Iwasa, Univ. Utah;绘图 @Nik Spencer/Nature SARS病毒会利用两个宿主蛋白酶中的一个入胞:TMPRSS2 或 组织蛋白酶L。
TMPRSS2的入胞途径更快,
但 SARS病毒经常通过核内体 (一种脂质包裹的囊泡)
侵入 ,这个途径
依赖组织蛋白酶L 。不过,如果病毒通过这一途径进入,就会被
抗病毒蛋白 【逮捕】。
新冠病毒之所以不同于 SARS病毒,就在于它能有效地利用 TMPRSS2 。
TMPRSS2是呼吸道细胞表面大量存在的一种酶,它会先
酶切刺突蛋白 S2亚基上的一个位点 。这个剪切点会暴露一串疏水性氨基酸,暴露后的疏水性氨基酸会迅速嵌入最近的膜中——即宿主细胞膜。
随后,
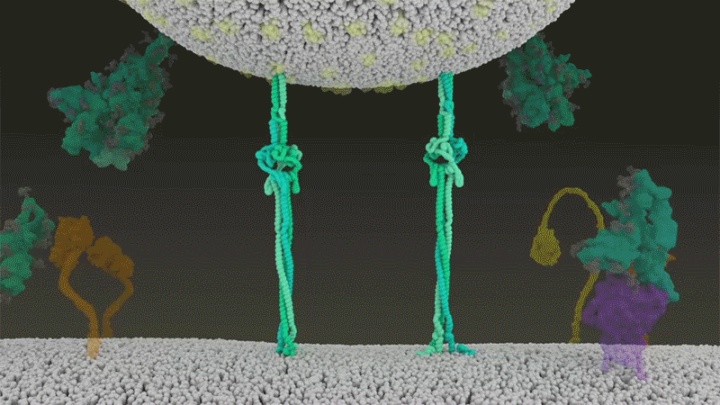
展开的刺突蛋白会折叠起来,像拉链一样,迫使病毒外膜与细胞膜融合。
新冠病毒与细胞融合过程的动画展示。来源 @Janet Iwasa, University of Utah
随后,
新冠病毒将基因组直接喷射到宿主细胞内 。
通过采取这种弹簧加载的侵入方式,
新冠病毒比 SARS病毒的感染更快,并避免被困在核内体。
新冠病毒能利用 TMPRSS2实现快速入胞,解释了为何疟疾药物
氯喹 一开始在实验中表现良好,但在治疗新冠的临床试验中无效。
结果证明,氯喹利用的细胞完全依赖组织蛋白酶实现核内体侵入。而新冠病毒在人类气道中传播和复制时不会用到核内体,所以
氯喹这种核内体干扰药物在真实人体中效果就不大了。
这一发现还指出,
蛋白酶抑制剂是一种很有前景的治疗选择 ,可以防止病毒利用 TMPRSS2、组织蛋白酶L 或其他蛋白酶进入宿主细胞。
如一种在日本已批准用于治疗胰腺炎 TMPRSS2抑制剂 camostat mesylate。这种抑制剂能阻断病毒进入肺部细胞,但无法在初步临床试验中改善病人的预后。
步骤③ —— 铲除异己
就在新冠病毒将 RNA基因组注射到宿主细胞后,细胞质中的核糖体会将两个病毒 RNA片段翻译成
氨基酸长链 ,这些氨基酸长链再被切割成 16个蛋白质,包括许多参与 RNA合成的蛋白质。
随后会产生更多的 RNA,这些 RNA编码总共有 26个已知的病毒蛋白,包括用于制造新病毒颗粒的结构蛋白(比如刺突蛋白)和其他协助蛋白。
通过这种方式,病毒开始大量复制自身的信使 RNA。 但它还需要细胞机器来将这些 mRNA翻译成蛋白质。
而新冠病毒有很多种方式将细胞机器占为己用:
① 首先,新冠病毒会清除竞争对手 :
病毒蛋白 Nsp1是新冠病毒首先被翻译的蛋白之一,它会招募宿主蛋白来系统地切割所有没有病毒标签的细胞 mRNA。
② 其次,感染使细胞中的全部蛋白质翻译减少70%:
Nsp1再次成为罪魁祸首,这次它物理阻断了核糖体的入胞通道,让 mRNA无法进入 ,剩余的少量翻译能力专用于翻译病毒 RNA。
③ 最后,新冠病毒关闭了细胞的预警系统:
研究团队发现了新冠病毒的一个明确机制:病毒让细胞 mRNA无法离开细胞核,包括用于提醒免疫系统感染的蛋白质指令。 因为 Nsp1蛋白似乎阻塞了离开细胞核的通道,因此没有任何东西可以逃脱。
由于基因转录本无法离开细胞核,因此受感染的细胞不会释放许多干扰素——
干扰素是提醒免疫系统注意病毒的信号蛋白。
新冠病毒在关闭此预警系统的方面特别高效:和其他呼吸道病毒相比(包括 SARS病毒和呼吸道合胞病毒),
新冠病毒感染后诱导的干扰素水平显著降低 。
今年6月,研究人员报告了Alpha变异株的突变似乎能更有效地抑制干扰素的产生 。
很明显,新冠病毒是一个动作非常快的病毒,具有独特的能力,可以从源头上扰乱免疫系统识别病毒和抵抗感染的能力。
当免疫系统真正发现存在病毒时,病毒数量已经太多了,以至于免疫应答蛋白有时会以平时更快的速度涌入血液,反而对人体有害。
医生在疫情早期就发现,
一些新冠重症患者会同时受到病毒本身及过度活跃的免疫应答的伤害 (一些行之有效的治疗方法就可以专门抑制这种免疫应答)
。
步骤④ —— 超强改造 当新冠病毒接管宿主的细胞翻译后,它就开始大肆改造了,根据它的需要广泛改造细胞的内部和表面。
首先,
一些新制造的刺突蛋白会移动到细胞表面并突破宿主细胞膜。 它们还会在那里
激活宿主钙离子通道,在细胞表面分泌一层脂肪膜 ——肌肉细胞等自然融合的细胞上也会发现这种膜。
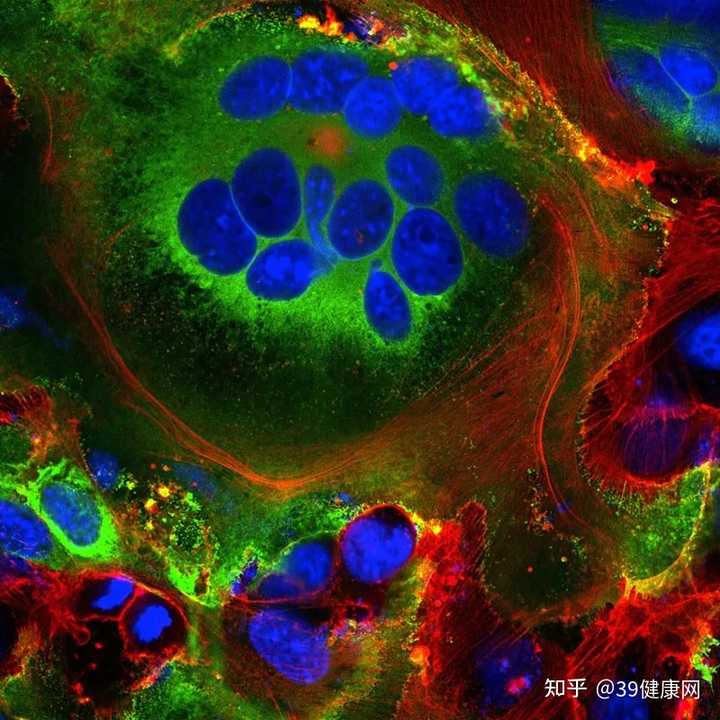
这时,受感染的细胞与表达 ACE2的相邻细胞融合,发育成
多达 20个细胞核的单个大呼吸道细胞。
这些融合结构被称为合胞体(syncytia),
HIV和单纯疱疹病毒等病毒感染就会引起,SARS病毒则不会 。
表达新冠病毒刺突蛋白(绿色)的细胞内可见融合细胞结构(合胞体)。蓝色为细胞核,红色为细胞骨架。来源 @Mauro Giacca
而研究结果表明,
合胞体的形成能让受感染的细胞存活更长时间,产生出更多的病毒颗粒。 也就是说新冠不是一个肇事逃逸的病毒,而是
会持续存在 。
中国医学科学院团队发现:
一些感染新冠病毒的细胞甚至会与淋巴细胞 (人体自身的免疫细胞之一)
形成合胞体 。
这其实是肿瘤细胞的免疫逃逸机制,而非病毒的机制,这也表明
受感染的细胞会通过抓住周围的免疫细胞并与之融合,从而轻而易举地逃过免疫系统的侦查。
细胞内部更多的变化在发生。与其他冠状病毒一样,
新冠病毒将细长的内质网 (ER) (一种参与蛋白质合成和运输的扁平膜网络)
转化为双膜球 ,就好像 ER在吹泡泡一样。
这些双膜囊泡 (DMV) 可能为病毒 RNA 的复制和翻译提供了一个安全的场所,使其免受细胞中先天免疫传感器的影响 (但该假设仍在研究中)。
大多数具有外层膜(即包膜)的病毒获得这一特征的方式是直接在细胞边缘组装,并在离开的路上选择一些细胞自身的质膜。
但是新制造的冠状病毒蛋白会选择走另一条途径。
虽然多年来的证据表明:
冠状病毒是通过高尔基体复合体运输出细胞的。
高尔基复合体是具有邮局功能的细胞器,能将分子包在膜内并运送至细胞其他部位。病毒会从高尔基体膜形成一个脂质包膜;然后新形成的病毒颗粒再通过高尔基囊泡转运到细胞表面,并被分泌出细胞。
但在去年 12月发现:
冠状病毒会通过溶酶体离开细胞。
溶酶体是细胞的垃圾桶,拥有很多能分解细胞成分的酶。
阻断基于高尔基体的分泌途径似乎不会影响释放的感染性病毒的数量。研究证据显示,
病毒蛋白通过进入内质网形成包膜,再占用溶酶体离开细胞 。
研究人员目前正在测试阻断溶酶体退出过程的抑制剂 作为潜在的抗病毒候选药物。
与从质膜中出芽相比,让细胞通过高尔基体或溶酶体是缓慢且低效的。所以
科学家们不明白新冠病毒为什么会这么做。
科学家推测,相比质膜,来自高尔基体或溶酶体的包膜的脂质组成好像对新冠病毒更有利。
步骤⑤ —— 攻城略地
在离开细胞的过程中,还有关键一招让新冠病毒成为传染王:
在有 5个氨基酸的位点进行快速切割,为病毒攻击下一个目标做好了准备。
其他冠状病毒在刺突蛋白 S1和 S2亚基的连接处只具有一个精氨酸,而
新冠病毒却具有 5个连着的氨基酸 :脯氨酸、精氨酸、精氨酸、丙氨酸、精氨酸。
这个位点不同寻常,确实是侵入肺部细胞的关键。
2020年 5月发现,这个名为
弗林蛋白酶 的宿主细胞蛋白能识别并切割这串氨基酸链,而且这种切割对于新冠病毒快速进入人类肺部细胞至关重要。
这就是在新冠病毒上发现的
弗林切割位点 (致病性很高的
禽流感病毒 也有这个位点)。
荷兰一项研究也发现,具有完整弗林切割位点使冠状病毒进入人体呼吸道细胞的速度更快。
研究人员推测弗林会在病毒颗粒组装过程中或是释放前切割该位点。这个时间点也解释了新冠病毒为何会通过高尔基体或溶酶体离开细胞。因为新冠病毒
一旦组装好就会移动到周围都是弗林蛋白酶的细胞器中。
通过剪切 S1和 S2亚基之间的键,弗林蛋白酶的切割松开了病毒颗粒的刺突蛋白,因此在进入细胞时,它们对 TMPRSS2的二次切割产生反应,这次切割暴露出的疏水性区域会快速将自己嵌入宿主细胞膜内。
如果刺突蛋白没有被弗林蛋白酶预先切开——有时也不会被切开——它们就会绕开 TMPRSS2,并通过较慢的核内体途径进入细胞。
而 Alpha和 Delta变异株的弗林切割位点都发生了变化。 Alpha变异株将本来的脯氨酸替换成组氨酸(P681H);Delta变异株则替换成了精氨酸(P681R)。这两个变化都会降低序列的酸性;而且氨基酸链碱性越强,被弗林识别切割的效率就越高。
这也导致了新冠病毒的传递能力增强。
弗林蛋白酶切割意味着更多刺突蛋白准备进入人体细胞。 SARS病毒只有不到 10%的刺突蛋白做好了这种准备,但对新冠病毒来说,这个比例上升至 50%,而 Alpha毒株超过了 50%。
研究发现:
在传播性更强的 Delta毒株中, 超过 75%以上的刺突蛋白已准备好感染人体细胞。
其实, 科学界对新冠病毒的了解还只是冰山一角,
一些关键的未知数还包括 :
与每个刺突蛋白结合所需的 ACE2受体数量;
S2位点究竟是何时被TMPRSS2切割的;
病毒外膜与细胞膜融合所需的刺突蛋白数量……
而这些还只是入胞的问题。
2020年 4月,加州大学旧金山分校的一个团队鉴定出了
新冠病毒与人类蛋白质之间至少有 332种相互作用。
想要赶上快速变异的病毒的步伐并不容易。
但专家一致认为,迄今为止,
大多数突变都与病毒传播的效率有关,而不是病毒对宿主的破坏程度。
7月的一项研究报道,与早期的病毒相比,
Delta变异株在肺部和喉部的生长速度更快,水平也更高 。
但是,
尚不确定 Delta的突变是如何以这种方式增强自身的 ,许多问题仍待解答。
原文标题:《How the coronavirus infects cells — and why Delta is so dangerous》
原作者:Megan Scudellari
原出处:《Nature》
原文链接:https://www. nature.com/articles/d41 586-021-02039-y